
SSE#208: ATUALIZAÇÃO SOBRE A SUPLEMENTAÇÃO COM BETA-ALANINA EM ATLETAS
PONTOS-CHAVE:
- A fadiga durante ~1-10 minutos nos esportes/eventos de alta intensidade é multifatorial, mas há fortes fundamentos de mecanismos demonstrando que a acidose da musculatura esquelética, através do acúmulo do íon de Hidrogênio (H+), é a chave para a limitação da performance. Assim, a musculatura esquelética tem vários mecanismos inatos de proteção intra e extracelulares para abordar a acidose induzida pelo exercício.
- A carnosina é um tampão intracelular importante devido ao seu conteúdo de nitrogênio com anel imidazol, que consegue aceitar (tamponar) H+, reduzir a velocidade no declínio do pH muscular durante exercícios intensos e contribuir com ~15% da capacidade total de tamponamento.
- Além do papel de tampão, a carnosina também foi mostrada participando da troca do cálcio difusível (Ca2+)/H+, fornecendo Ca2+ de volta ao retículo endoplasmático e H+ para a membrana celular, sugerindo que isso pode melhorar a sensibilidade muscular ao Ca2+ e a eficiência da contração muscular.
- A carnosina da musculatura é sintetizada pela carnosina sintase, processo do qual a concentração plasmática de beta-alanina é o substrato limitante; dados consistentes demonstraram que a suplementação com ~3-6g de beta-alanina por pelo menos 4 semanas pode aumentar o estoque muscular de carnosina em 30-60%.
- Diversas meta-análises mostraram tamanho de efeito moderado para a capacidade de exercício, e menor tamanho de efeito para a performance em exercícios com ~1-10 minutos de duração. Isto se traduz em ~2-3% de benefícios na performance em indivíduos ativos que não pertencem ao grupo de elite, mas ~0,5-1% de aumento na performance em indivíduos do grupo de elite. No entanto, mais dados são necessários para populações de atletas de elite.
- Apesar do considerável aumento no conhecimento científico em relação aos protocolos de suplementação com beta-alanina e sua eficácia na performance, desde 2006 há muitas questões determinantes que permanecem e a necessidade de um direcionamento para futuras pesquisas aplicadas.
INTRODUÇÃO
Atletas que participam em esportes/eventos de alta intensidade (~1-10 minutos com esforço total), ou em esportes onde os atletas necessitam fazer esforço repetitivo de alta intensidade, têm determinantes específicos de performance. Todos estes esportes utilizam grande quantidade de produção de energia do adenosina trifosfato (ATP) derivado anaerobicamente da fosfocreatina e da glicólise anaeróbica, a última resultando em acúmulo de grande quantidade de lactato (> 10 mmol/l). A diversidade destes determinantes específicos de performance de alta intensidade é especialmente prevalente em eventos de meia-distância/alta-intensidade que apresentam uma mistura de características aeróbicas, anaeróbicas e neuromusculares/mecânicas (Sandford & Stellingwerff, 2019), e muitos destes determinantes estão ligados a intervenções nutricionais (Stellingwerff, Bovim, & Whitfield, 2019). Certamente, os limites da performance de alta intensidade são multifatoriais, mas o principal fator limitante é a habilidade de tolerar níveis de acidose muscular em constante aumento, tanto intra como extracelular. Para melhorar o efeito tampão extracelular, a dose de bicarbonato de sódio foi pesquisada e utilizada por atletas por muitas décadas.
No entanto, foi apenas no meio da década de 2000 que um trabalho pioneiro do Professor Roger Harris e colaboradores (2006) demonstrou que aumentar o tamponamento intracelular (dentro do músculo) era também possível via suplementação crônica (várias semanas) com beta-alanina, que aumentava significativamente o conteúdo de carnosina muscular (b-alanil-L-histidina) e a performance de alta intensidade (Hill et al., 2007) (Figura 1). Desde esta época, houve uma explosão de pesquisas examinando a eficácia da suplementação com beta-alanina para aumentar de forma máxima o conteúdo muscular de carnosina e melhorar a performance como consequência. Portanto, este artigo do Sports Science Exchange (SSE) irá primeiro examinar as limitações para a performance de alta intensidade, com ênfase no metabolismo anaeróbico e os mecanismos de tamponamento associados, e então examinar a ciência dos protocolos de suplementação com beta-alanina e seus diversos resultados na performance.
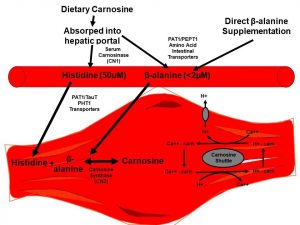
Figura 1: Esquema delineando a captação de beta-alanina e a consequente síntese, estoque e mecanismos propostos envolvendo o aumento do conteúdo de carnosina. β–alanina, beta-alanina; Carn, carnosina; Ca++, cálcio; CN1, carnosina plasmática; CN2, carnosina sintase; H+, íon de hidrogênio; PAT1, transportador de aminoácidos acoplado a prótons 1; PEPT1, transportador de peptídeos 1; PHT1, transportador de peptídeos histidina 1; Tau-T, transportador de taurina; Figura adaptada de (Blancquaert et al., 2015; Matthews et al., 2019; Stellingwerff, Decombaz, et al., 2012).
Tradução da figura:
Carnosina da dieta//Absorvida pela veia portal hepática
Suplementação direta com beta-alanina
Histidina
LIMITAÇÕES NA PERFORMANCE EM EXERCÍCIOS DE ALTA INTENSIDADE E OS MECANISMOS DE AÇÃO DA CARNOSINA MUSCULAR
Eventos/esportes de alta intensidade (~1-10 minutos de esforço total) são específicos já que eles se encontram em uma intersecção metabólica e recebem grandes contribuições de energia tanto do metabolismo aeróbico, quanto do anaeróbico (Figura 2). Existem três principais sistemas de energia que fornecem ATP simultaneamente (apesar de acontecerem em diferentes velocidades) para alimentar o exercício de alta intensidade, especialmente os sistemas anaeróbicos (também chamados de fosforilação ao nível de substrato): 1) degradação de fosfocreatina (PCr), 2) glicólise “anaeróbica”, e 3) metabolismo aeróbico (também conhecido como fosforilação oxidativa). Ao início do exercício e durante situações de exercício com intensidade progressiva, a produção de ATP do metabolismo aeróbico não pode igualar com a taxa de utilização de ATP (100% dos eventos de alta intensidade), e a carência de fornecimento de energia é compensada pelo metabolismo anaeróbico. Esta energia é fornecida pelos sistemas de PCr e glicólise anaeróbica. A última tem uma capacidade maior que a degradação de PCr e fornece principalmente ATP através da quebra de glicogênio na via glicolítica, resultando em produção excessiva de piruvato. Durante exercícios extremamente intensos, a alta taxa de produção de piruvato excede a velocidade com que ele pode ser oxidado aerobicamente pela piruvato desidrogenase na mitocôndria, através do ciclo do ácido tricarboxílico e da cadeia transportadora de elétrons (Spriet, Howlett, & Heigenhauser, 2000), levando a produção de níveis extremos de lactato associada aos esportes de alta intensidade (competições e treinos).
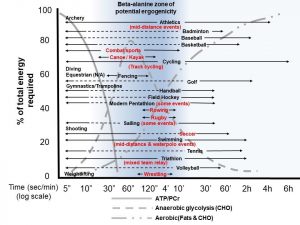
Figura 2: Esquema destacando a zona de potencial efeito ergogênico da beta-alanina (~1-10 minutos de exercício de alta intensidade) nos esportes olímpicos. Cada um dos três principais sistemas de energia de produção de ATP é destacado ao fundo, demonstrando a sobreposição considerável que acontece nos esportes/eventos que têm grande dependência da glicólise anaeróbica, e a possibilidade de melhorar a performance com a suplementação de beta-alanina. Os esportes/eventos destacados em vermelho apresentam a existência de dados publicados sobre os níveis de lactato maiores que 10 mmol/l em competições. A duração total do evento/esporte é destacada pela flecha a direita. A linha pontilhada destaca descontinuidade na duração (ou a alta intensidade não mantida (< 0,5 minutos)) em cada esporte (exemplo, sprints curtos com recuperação aeróbica prolongada onde os níveis de lactato são < 10mmol/l, como durante jogos de futebol ou baseball). ATP, trifosfato de adenosina; CHO, carboidrato; log, logaritmo; mid-distance, meia-distância; min, minuto; PCr, fosfocreatina; sec, segundos).
Tradução da figura:
Zona de Potencial Ergogênico da Beta-Alanina
% da Energia Necessária Total
Tempo (seg/min)
Escala logarítmica
Glicólise anaeróbica (CHO)
Aeróbica (Gorduras & CHO)
Arco e flecha
Atletismo/(eventos de meia-distância)
Badminton
Baseball
Basquete
Esportes de combate
Canoagem/caiaque
Ciclismo
(ciclismo em pista)
Mergulho
Hipismo (N/A)
Esgrima
Golfe
Ginástica/Trampolim
Handball
Hockey de campo
Pentatlo moderno (alguns eventos)
Remo
Rugby
Barco à vela (alguns eventos)
Tiro
Futebol
Natação
(eventos de meia-distância e polo aquático)
Tênis
Triathlon
(revezamento misto)
Vôlei
Levantamento de peso
Luta livre
Desta forma, o esforço de alta intensidade com duração ~4 minutos é conduzido em ~20 vezes os valores do VO2 de repouso, resultando em níveis de lactato sanguíneo tão altos quanto ~25 mmol/l, e a produção de íons de hidrogênio causando redução no pH da musculatura esquelética humana de ~7,2 em repouso para 6,6 na fadiga (Hermansen & Osnes, 1972). Apesar da fadiga ser multifatorial, há certamente um forte fundamento de mecanismos demonstrando a acidose muscular, através do acúmulo de H+, como um importante limitante para a manutenção do exercício de alta intensidade por ~1-10 minutos (Allen, Lamb, & Westerblad, 2008). Também é conhecido por mais de 80 anos que a carnosina é um tampão intracelular essencial devido ao seu nitrogênio com anel imidazol, que pode aceitar (tamponar) H+ (Bate-Smith, 1938), e reduzir a velocidade do declínio do pH muscular durante exercícios intensos (Baguet, Koppo, Pottier, & Derave, 2010). A contribuição dos níveis normais da carnosina muscular para a capacidade completa de tamponamento intracelular muscular foi sugerida ser de ~6-7%, mas pode atingir ~15% do total quando elevado através da suplementação com beta-alanina (Harris & Stellingwerff, 2013). Curiosamente, também é conhecido por 35 anos que velocistas e atletas do remo têm quase o dobro da quantidade de carnosina muscular que corredores de maratona, e que a carnosina se correlaciona fortemente com o tipo II de fibra muscular (Parkhouse, McKenzie, Hochachka, & Ovalle, 1985). De maneira global, a acidose metabólica severa é um limitante para a performance de alta intensidade, na qual a carnosina muscular pode agir como um dos muitos tampões intracelulares.
Além da acidose metabólica limitar a produção de ATP, há também restrições biomecânicas e estruturais na performance de alta intensidade que frequentemente não são consideradas quando se esclarece os limites para a performance de alta intensidade. Por exemplo, muitos esportes de meia-distância/de força necessitam de adaptações morfológicas extremas (exemplo, circunferência excepcional dos quadríceps na corrida de ciclismo e remo) que resultam em velocidade e força explosiva e/ou energia extraordinárias. Obviamente os determinantes de performance destes esportes também precisam ser considerados a partir de uma origem estrutural e biomecânica (exemplo, pico de força absoluto, taxa de desenvolvimento de força, massa muscular etc. (Weyand, Sandell, Prime, & Bundle, 2010)). A corrida de velocidade de alta intensidade sustentada próximo da atividade máxima parece ser limitada pelo mínimo de tempo necessário (tempo de contato inicial ou tempo sob pressão) para aplicação de grandes forças, que são novamente relacionadas com o conteúdo de fibras musculares do tipo II de contração rápida (Weyand et al., 2010) e/ou com a habilidade da musculatura em repetidamente gerar força máxima. Sustentando estes diversos determinantes estruturais/mecânicos de performance estão as fibras musculares do tipo II de contração rápida (Tipo IIa e IIx) (Costill et al., 1976)), que têm quase o dobro de conteúdo de carnosina muscular em comparação com fibras musculares do Tipo I de contração lenta (Baguet et al., 2011; Parkhouse et al., 1985; Stellingwerff, Anwander, et al., 2012).
A carnosina também tem inúmeros papéis fisiológicos, apesar da maioria do fundamento destes mecanismos ter sido gerado através de pesquisas com roedores e culturas celulares. Contudo, muitos destes papéis in vitro podem estar agindo durante o repouso e nos exercícios em humanos também. Todos estes possíveis papéis fisiológicos foram revisados recentemente de maneira significativa por Matthews et al. (2019), com a melhora no tamponamento do pH intracelular sendo o mecanismo ergogênico mais fortemente predominante em relação ao maior conteúdo de carnosina em atletas. Os mecanismos secundários também incluem: (1) o manejo do cálcio (Ca2+), incluindo liberação, recaptação e sensibilidade ao Ca2+; (2) troca de Ca2+/ H+ citoplasmática descrita como o “transporte da carnosina” (Figura 1); (3) possibilidade de regular a bioenergética, levando a um aumento no fluxo glicolítico; (4) remoção das espécies reativas de oxigênio, alterando o estresse oxidativo; e (5) formação de compostos estáveis para prevenir a ocorrência de aldeídos reativos e peroxidação lipídica (detoxificação não-enzimática dos aldeídos reativos).
Um recente mecanismo interessante é a hipótese do transporte de carnosina (Figura 1). Como suporte para esta hipótese do transporte da carnosina, dados recentes sobre os miócitos cardíacos demonstraram que a carnosina não é apenas um tampão, mas também está envolvida no manejo do Ca2+ e H+ do retículo endoplasmático como um tipo de “transporte de carnosina” (Figura 1; como revisado por (Blancquaert, Everaert, & Derave, 2015; Matthews, Artioli, Turner, & Sale, 2019)), mas isso ainda necessita de confirmação na musculatura esquelética humana.
SUPLEMENTAÇÃO COM BETA-ALANINA E A SÍNTESE DE CARNOSINA
A carnosina da musculatura é sintetizada pela carnosina sintase, processo no qual a concentração plasmática de beta-alanina é o substrato limitante. A concentração plasmática é < 2 µM e o valor da Km (concentração necessária para a velocidade da reação que é 50% da máxima) para sua captação na musculatura é de ~1,0-2,3 mM. Ao contrário, a L-histidina está presente em concentrações muito maiores no plasma (50 µM) e na musculatura, e tem uma Km muito mais baixa (16,8 µM, Figura 1). Em 2006, Harris e colaboradores (2006) foram os primeiros a demonstrar que a ingestão oral de 3,2 ou 6,4 g/dia de beta-alanina (ou uma quantidade isomolar da carnosina suplementada (13,0 g/dia)), resultou em um aumento de 40-60% da carnosina muscular em humanos.
Desde este estudo de referência, cada estudo que mediu a carnosina muscular (seja através de biópsia muscular ou espectroscopia por ressonância magnética), onde uma quantidade significativa de beta-alanina foi suplementada (exemplo ~3-6g de beta-alanina/dia por pelo menos 4 semanas), mostrou um aumento significativo do conteúdo de carnosina muscular (para revisões veja (Perim et al., 2019; Stellingwerff, Decombaz, Harris, & Boesch, 2012). Em média, isto levou a um aumento considerável ~30-50% no conteúdo de carnosina muscular com suplementação com ~3-6g/dia de beta-alanina por 4-6 semanas. O “washout” da carnosina com conteúdo aumentado na musculatura esquelética após o término da suplementação com beta-alanina é muito lento, com um tempo estimado de washout de ~14 a 15 semanas após um aumento de ~50% na carnosina muscular (Baguet et al., 2009; Stellingwerff, Anwander, et al., 2012). De longe o maior fator contribuinte para a síntese de carnosina é a dose e a duração da suplementação com beta-alanina, já que estudos mostram que dobrar a dose diária de beta-alanina resulta em aproximadamente o dobro da quantidade de carnosina sintetizada (e metade do tempo para atingir o mesmo aumento) (Church et al., 2017; Stellingwerff, Anwander, et al., 2012).
Dados atuais também mostraram diversas possíveis maneiras de se aumentar a eficiência da síntese de carnosina com a suplementação com beta-alanina, já que apenas ~5% da beta-alanina ingerida pode ser contabilizada na musculatura como carnosina , com o restante tendo um destino metabólico desconhecido. Estas maneiras incluem: (1) consumir beta-alanina com a refeição (fornecendo um efeito insulínico), já que isto resultou em maior síntese de carnosina no músculo sóleo, mas não no músculo gastrocnêmio (Stegen et al., 2013); (2) utilizar a beta-alanina de liberação lenta em contraste a sua forma pura, resulta em maior síntese de carnosina (Varanoske et al., 2019); e (3) realização de exercícios/treinos durante a fase de suplementação (Bex, Chung, Baguet, Achten, & Derave, 2015). No entanto, além da dose e duração da suplementação com beta-alanina para impactar a síntese de carnosina, estes outros fatores de influência necessitam de mais validação científica. Além disso, um estudo de referência recente de Saunders et al. (2017a) que implementou 24 semanas de suplementação com beta-alanina com 6,4 g/dia em 25 indivíduos, relatou uma enorme diversidade na variabilidade individual para a síntese de carnosina. Cinco indivíduos tiveram conteúdos máximos de carnosina muscular em 24 semanas, mas outros 5 indivíduos já atingiram seus picos no conteúdo muscular de carnosina nas primeiras 12 semanas (Saunders, Painelli, et al., 2017). Finalmente, parece que uma vez que a carnosina muscular é elevada com uma fase de carga de beta-alanina, a continuação da suplementação com apenas 1,2 g/dia de beta-alanina mantém os estoques de carnosina elevados (Stegen et al., 2014). Obviamente, nosso entendimento dos determinantes da resposta individual para a suplementação de beta-alanina estão longe de serem completamente compreendidos (Tabela 1).

Tabela 1: Questões remanescentes e direcionamento para futuras pesquisas aplicadas examinando a eficácia da suplementação com beta-alanina na melhora da performance de alta intensidade e/ou adaptação aos treinos.
| Beta-alanina: questões remanescentes e direcionamento para futuras pesquisas aplicadas |
| Quais são as razões mecanicistas, além do valor de referência da carnosina, para as respostas de indivíduos maiores à síntese de carnosina a partir da suplementação com beta-alanina (Saunders, Painelli, et al., 2017)? |
| Já que a carnosina é dependente do tipo de fibra muscular (Baguet et al., 2011), há respostas específicas do tipo de fibra muscular em relação aos resultados de performance ou potencial ergogênico da beta-alanina baseando-se no conteúdo de referência da carnosina (e portanto, do tipo de fibra)? |
| Há alguma implicação (exemplo, atenuação enzimática, protéica ou do transportador) da suplementação a longo-prazo com beta-alanina? (> 24 semanas, (Saunders, Painelli, et al., 2017)) |
| A hipótese de transporte de carnosina (Figura 1), como principalmente desenvolvido em modelos de cultura celular, também ocorre em humanos in vivo via aumento de H+ e/ou manejo do cálcio Ca2+? Como estes mecanismos separados, porém relacionados, impactam nos resultados de performance em humanos? |
| Mais estudos sobre treinos, examinando a eficácia em relação à adaptação aos treinos da suplementação com beta-alanina levando ao aumento na carnosina, são necessários – a maior carga de atividade causa maior adaptação aos treinos ou uma possível atenuação do H+/acidose impacta na adaptação? |
| Já que a avaliação da carnosina na musculatura esquelética humana realizada pela espectroscopia por ressonância magnética por próton não-invasiva pode ser utilizada em atletas para diferenciar o tipo de fibra (Baguet et al., 2011), há diferenças distintas em relação ao tipo de fibra em todos os aspectos do treinamento e recuperação? Como a suplementação com beta-alanina pode impactar nestes parâmetros? |
| Há alguma alteração na avaliação do ritmo (teleantecipação) que ocorre quando se altera o conteúdo de carnosina muscular que necessite de consideração cautelosa em relação aos efeitos nos testes de tempo como no ciclismo, natação e remo? |
Quando a beta-alanina é suplementada em quantidade maior que 800 mg/dose ela resulta em parestesia leve na maioria dos indivíduos (leve dormência e formigamento, vasodilatação da pele e vermelhidão por ~60-120 minutos). Contudo, um tablete de beta-alanina de liberação lenta comercializado tem sido produzido e demonstrou menor extravasamento urinário e curva menos acentuada do pico plasmático (a mesma área abaixo da curva), com nenhum dos indivíduos relatando qualquer sintoma de parestesia em comparação com o controle (Decombaz, Beaumont, Vuichoud, Bouisset, & Stellingwerff, 2012). Além da menor expressão do gene transportador de beta-alanina muscular e da proteína Tau-T (Saunders, Painelli, et al., 2017), nenhum outro efeito colateral ou alterações em quaisquer medidas sanguíneas, ou mudanças agudas de peso corporal foram demonstrados com a suplementação de beta-alanina (Harris et al., 2006; Stellingwerff, Anwander, et al., 2012).
RESULTADOS DE PERFORMANCE APÓS AUMENTO DA CARNOSINA PELA SUPLEMENTAÇÃO COM BETA-ALANINA
O primeiro estudo sobre a beta-alanina na performance humana foi publicado em 2007 e demonstrou que um aumento na carnosina muscular (pela suplementação com beta-alanina) resultou em aumentos dose-resposta no tempo total de exaustão (TTE) na atividade realizada em 110% da potência máxima (Wmax), em 4 semanas (+12% TTE) e em 10 semanas (+16% TTE), em comparação com a base de referência (Hill et al., 2007). Nos anos seguintes, uma infinidade de estudos baseados na performance demonstraram a eficácia da suplementação com beta-alanina na melhora dos resultados em atividades de alta intensidade, incluindo: (1) a carnosina muscular de referência pré-suplementação sendo correlacionada com ~16 segundos, ~1,5 minutos, e ~6,5 minutos na performance de remo (Baguet, Bourgois, Vanhee, Achten, & Derave, 2010), mostrando a importância da carnosina muscular na performance em exercícios de alta intensidade; (2) melhorou significativamente a performance em sprint de 30 segundos (+5% da força média) após atividade extensa de ciclismo com duração de 110 minutos (Van Thienen et al., 2009); e (3) melhores resultados em teste de Wingate de repetição (total de atividades realizadas em 4 testes de Wingate), tanto em indivíduos treinados como não-treinados (de Salles Painelli et al., 2014). No entanto, alguns estudos não demonstraram benefícios na performance após a suplementação com beta-alanina em comparação com o controle, mas de maneira geral nestes estudos, ou a suplementação foi menor que ~150g do total da suplementação com beta-alanina (Derave et al., 2007; Smith et al., 2009), ou os protocolos de performance foram muito curtos e explosivos, abaixo de ~30 segundos de duração (1 repetição ou 1 corrida, no máximo), casos em que a acidose não é considerada um fator limitante (Kendrick et al., 2008; Sweeney, Wright, Glenn Brice, & Doberstein, 2010).
Houve duas meta-análises decisivas que examinaram os efeitos na performance com a suplementação crônica de beta-alanina. A primeira meta-análise foi publicada em 2012 e destacou 15 estudos (n=360 indivíduos), mostrando uma conclusão geral da beta-alanina melhorando as medidas resultantes dos exercícios em comparação com o controle (p=0,002 com tamanho de efeito (ES) geral de 0,374), e a maioria dos efeitos de performance sendo demonstrados ao longo de 1 a 4 minutos de exercícios com alta intensidade (Hobson, Saunders, Ball, Harris, & Sale, 2012). Contudo, esta meta-análise foi dominada por protocolos de capacidade de exercício/TTE (n=13 estudos; p=0,013, ~2-3% de melhora) em comparação com protocolos de performance do exercício/teste de tempo (TT) (n=2 estudos; p=0,204). É importante notar que o tamanho de efeito tende a ser maior em protocolos TTE baseados na duração da atividade, em comparação com protocolos TT onde o ritmo intrínseco da atividade pode também ter um papel nos resultados da performance. Uma meta-análise de acompanhamento foi finalizada em 2017 mostrando 40 estudos (n=1461 indivíduos), onde o tamanho de efeito geral foi 2,1 vezes menor (ES=0,18) em comparação com a meta-análise de 2012 (Saunders, Elliott-Sale, et al., 2017). Curiosamente, o ES para a capacidade de exercício/TTE foi de 0,4998, similar à meta-análise de 2012, mas os resultados do ES para a performance/TT foi de apenas 0,1078. Novamente, a meta-análise com modelo de regressão mostrou que a duração do exercício de ~1-10 minutos demonstrou os melhores benefícios de performance com a suplementação com beta-alanina (p=0,004; Figura 2).
Finalmente, houve alguns dados recentes sugerindo efeitos menores na performance de maneira geral (~0,5-1% de melhora na performance) com a suplementação com beta-alanina em indivíduos bem treinados em comparação a indivíduos não-treinados (Bellinger, Howe, Shing, & Fell, 2012; Saunders, Elliott-Sale, et al., 2017). Isto foi demonstrado em outras meta-análises no passado e provavelmente representa o efeito biológico máximo em relação à performance, encontrado em atletas de elite atingindo o seu auge genético. A Figura 2 destaca os diversos sistemas de energia e o “ponto ideal” da performance em atividades com ~1-10 minutos de duração, e os diversos esportes olímpicos que desta forma podem se beneficiar com a suplementação com beta-alanina. A beta-alanina é um suplemento, dentre alguns, com suporte ergogênico a serem listados como possivelmente ergogênicos (quando utilizados com o protocolo correto em esportes/eventos indicados) como parte do recente consenso sobre suplementação do Comitê Olímpico Internacional (Peeling, Binnie, Goods, Sim, & Burke, 2018).
APLICAÇÕES PRÁTICAS E CONCLUSÕES
Desde que Harris e colaboradores mostraram primeiramente em 2006 que a suplementação crônica com beta-alanina aumentou significativamente o conteúdo muscular de carnosina em humanos, houve um grande avanço no nosso conhecimento em relação aos protocolos de suplementação e consequentes resultados na performance humana. Contudo, a Tabela 1 destaca algumas das questões remanescentes e um direcionamento para futuras pesquisas aplicadas.
Além destas limitações pendentes no nosso entendimento, as aplicações práticas e as recomendações atuais para a suplementação com a beta-alanina são:
- Os dados respaldam o consumo de ~3-6 g/dia de beta-alanina (geralmente doses de 800mg a 1,6g, a cada 4 horas com as refeições), por pelo menos 4 semanas de suplementação, para aumentar os estoques de carnosina (30-60%) e apresentar benefícios na performance.
- Há evidências de que esta dosagem pode ser otimizada em relação ao peso corporal (~0,05-0,06 g/kg de massa corporal/dia), e que a beta-alanina de liberação lenta, ingerida com alimentos e durante fase de exercícios, irá melhorar a síntese adicional de carnosina (Stegen et al., 2014). Contudo, com 24 semanas de suplementação alguns atletas podem ainda estar aumentando seu conteúdo muscular de carnosina.
- Duas meta-análises mostraram que esportes/eventos de alta intensidade (~1-10 minutos de duração, com níveis de lactato sanguíneo significativamente altos (>10 mmol/l, Figura 2)) parecem ter maior potencial ergogênico com a suplementação com beta-alanina.
- Efeitos pequenos a moderados na performance foram relatados com a utilização da beta-alanina em eventos que mantêm a intensidade alta, com 2-3% de aumento na performance em atletas recreacionais, e ~0,5-1,0 em atletas de elite.
Este SSE fornece uma atualização sobre a ciência aplicada relacionada a suplementação com beta-alanina e seu impacto no conteúdo de carnosina muscular, e resultados de performance no exercício. Apesar de muitas questões remanescentes (Tabela 1), a explosão de conhecimento nos últimos 15 anos fez da beta-alanina um suplemento com potencial ergogênico dentre alguns suplementos, quando utilizada com protocolo adequado de dosagem, em eventos que mantêm a intensidade alta.
Os pontos de vista expressos são dos autores e não necessariamente refletem a posição ou política da PepsiCo, Inc.
REFERÊNCIAS
Allen, D. G., Lamb, G. D., & Westerblad, H. (2008). Skeletal muscle fatigue: cellular mechanisms. Physiological reviews, 88(1), 287-332. doi:10.1152/physrev.00015.2007
Baguet, A., Bourgois, J., Vanhee, L., Achten, E., & Derave, W. (2010). Important role of muscle carnosine in rowing performance. Journal of Applied Physiology, 109(4), 1096-1101. doi:10.1152/japplphysiol.00141.2010
Baguet, A., Everaert, I., Hespel, P., Petrovic, M., Achten, E., & Derave, W. (2011). A new method for non-invasive estimation of human muscle fiber type composition. PLoS ONE, 6(7), e21956. doi:10.1371/journal.pone.0021956
Baguet, A., Koppo, K., Pottier, A., & Derave, W. (2010). Beta-alanine supplementation reduces acidosis but not oxygen uptake response during high-intensity cycling exercise. European Journal of Applied Physiology, 108(3), 495-503. doi:10.1007/s00421-009-1225-0
Baguet, A., Reyngoudt, H., Pottier, A., Everaert, I., Callens, S., Achten, E., & Derave, W. (2009). Carnosine loading and washout in human skeletal muscles. Journal of Applied Physiology, 106(3), 837-842. doi:10.1152/japplphysiol.91357.2008
Bate-Smith, E. C. (1938). The buffering of muscle in rigor: protein, phosphate and carnosine. Journal of Physiology, 92(3), 336-343.
Bellinger, P. M., Howe, S. T., Shing, C. M., & Fell, J. W. (2012). Effect of combined beta-alanine and sodium bicarbonate supplementation on cycling performance. Medicine and science in sports and exercise, 44(8), 1545-1551. doi:10.1249/MSS.0b013e31824cc08d
Bex, T., Chung, W., Baguet, A., Achten, E., & Derave, W. (2015). Exercise training and Beta-alanine-induced muscle carnosine loading. Front Nutr, 2, 13. doi:10.3389/fnut.2015.00013
Blancquaert, L., Everaert, I., & Derave, W. (2015). Beta-alanine supplementation, muscle carnosine and exercise performance. Current opinion in clinical nutrition and metabolic care, 18(1), 63-70. doi:10.1097/MCO.0000000000000127
Church, D. D., Hoffman, J. R., Varanoske, A. N., Wang, R., Baker, K. M., La Monica, M. B., . . . Stout, J. R. (2017). Comparison of Two beta-Alanine Dosing Protocols on Muscle Carnosine Elevations. J Am Coll Nutr, 36(8), 608-616. doi:10.1080/07315724.2017.1335250
Costill, D. L., Daniels, J., Evans, W., Fink, W., Krahenbuhl, G., & Saltin, B. (1976). Skeletal muscle enzymes and fiber composition in male and female track athletes. Journal of Applied Physiology, 40(2), 149-154. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/129449
de Salles Painelli, V., Saunders, B., Sale, C., Harris, R. C., Solis, M. Y., Roschel, H., . . . Lancha, A. H., Jr. (2014). Influence of training status on high-intensity intermittent performance in response to beta-alanine supplementation. Amino Acids, 46(5), 1207-1215. doi:10.1007/s00726-014-1678-2
Decombaz, J., Beaumont, M., Vuichoud, J., Bouisset, F., & Stellingwerff, T. (2012). Effect of slow-release beta-alanine tablets on absorption kinetics and paresthesia. Amino Acids, 43(1), 67-76. doi:10.1007/s00726-011-1169-7
Derave, W., Ozdemir, M. S., Harris, R. C., Pottier, A., Reyngoudt, H., Koppo, K., . . . Achten, E. (2007). beta-Alanine supplementation augments muscle carnosine content and attenuates fatigue during repeated isokinetic contraction bouts in trained sprinters. Journal of Applied Physiology, 103(5), 1736-1743. Retrieved from http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=17690198
Harris, R. C., & Stellingwerff, T. (2013). Effect of beta-alanine supplementation on high-intensity exercise performance. Nestle Nutr Inst Workshop Ser, 76, 61-71. doi:10.1159/000350258
Harris, R. C., Tallon, M. J., Dunnett, M., Boobis, L., Coakley, J., Kim, H. J., . . . Wise, J. A. (2006). The absorption of orally supplied beta-alanine and its effect on muscle carnosine synthesis in human vastus lateralis. Amino Acids, 30(3), 279-289.
Hermansen, L., & Osnes, J. B. (1972). Blood and muscle pH after maximal exercise in man. Journal of Applied Physiology, 32(3), 304-308. doi:10.1152/jappl.1972.32.3.304
Hill, C. A., Harris, R. C., Kim, H. J., Harris, B. D., Sale, C., Boobis, L. H., . . . Wise, J. A. (2007). Influence of beta-alanine supplementation on skeletal muscle carnosine concentrations and high intensity cycling capacity. Amino Acids, 32(2), 225-233. Retrieved from http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=16868650
Hobson, R. M., Saunders, B., Ball, G., Harris, R. C., & Sale, C. (2012). Effects of beta-alanine supplementation on exercise performance: a meta-analysis. Amino Acids, 43(1), 25-37. doi:10.1007/s00726-011-1200-z
Kendrick, I. P., Harris, R. C., Kim, H. J., Kim, C. K., Dang, V. H., Lam, T. Q., . . . Wise, J. A. (2008). The effects of 10 weeks of resistance training combined with beta-alanine supplementation on whole body strength, force production, muscular endurance and body composition. Amino Acids, 34(4), 547-554. Retrieved from http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=18175046
Matthews, J. J., Artioli, G. G., Turner, M. D., & Sale, C. (2019). The Physiological Roles of Carnosine and beta-Alanine in Exercising Human Skeletal Muscle. Medicine and science in sports and exercise, 51(10), 2098-2108. doi:10.1249/MSS.0000000000002033
Parkhouse, W. S., McKenzie, D. C., Hochachka, P. W., & Ovalle, W. K. (1985). Buffering capacity of deproteinized human vastus lateralis muscle. Journal of Applied Physiology, 58(1), 14-17. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/3968004
Peeling, P., Binnie, M. J., Goods, P. S. R., Sim, M., & Burke, L. M. (2018). Evidence-Based Supplements for the Enhancement of Athletic Performance. Int J Sport Nutr Exerc Metab, 28(2), 178-187. doi:10.1123/ijsnem.2017-0343
Perim, P., Marticorena, F. M., Ribeiro, F., Barreto, G., Gobbi, N., Kerksick, C., . . . Saunders, B. (2019). Can the Skeletal Muscle Carnosine Response to Beta-Alanine Supplementation Be Optimized? Front Nutr, 6, 135. doi:10.3389/fnut.2019.00135
Sandford, G. N., & Stellingwerff, T. (2019). ‘Question your categories’: the misunderstood complexity of middle-distance running profiles with implications for research methods and application. Front. Sports Act. Living. doi:https://doi.org/10.3389/fspor.2019.00028
Saunders, B., Elliott-Sale, K., Artioli, G. G., Swinton, P. A., Dolan, E., Roschel, H., . . . Gualano, B. (2017). beta-alanine supplementation to improve exercise capacity and performance: a systematic review and meta-analysis. Br J Sports Med, 51(8), 658-669. doi:10.1136/bjsports-2016-096396
Saunders, B., Painelli, V. D. S., Oliveira, L. F., Silva, V. D. E., Silva, R. P., Riani, L., . . . Gualano, B. (2017). Twenty-four Weeks of beta-Alanine Supplementation on Carnosine Content, Related Genes, and Exercise. Medicine and science in sports and exercise, 49(5), 896-906. doi:10.1249/MSS.0000000000001173
Smith, A. E., Moon, J. R., Kendall, K. L., Graef, J. L., Lockwood, C. M., Walter, A. A., . . . Stout, J. R. (2009). The effects of beta-alanine supplementation and high-intensity interval training on neuromuscular fatigue and muscle function. European Journal of Applied Physiology, 105(3), 357-363. doi:10.1007/s00421-008-0911-7
Spriet, L. L., Howlett, R. A., & Heigenhauser, G. J. (2000). An enzymatic approach to lactate production in human skeletal muscle during exercise. Medicine and science in sports and exercise, 32(4), 756-763. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/10776894
Stegen, S., Bex, T., Vervaet, C., Vanhee, L., Achten, E., & Derave, W. (2014). beta-Alanine dose for maintaining moderately elevated muscle carnosine levels. Medicine and science in sports and exercise, 46(7), 1426-1432. doi:10.1249/MSS.0000000000000248
Stegen, S., Blancquaert, L., Everaert, I., Bex, T., Taes, Y., Calders, P., . . . Derave, W. (2013). Meal and beta-alanine coingestion enhances muscle carnosine loading. Medicine and science in sports and exercise, 45(8), 1478-1485. doi:10.1249/MSS.0b013e31828ab073
Stellingwerff, T., Anwander, H., Egger, A., Buehler, T., Kreis, R., Decombaz, J., & Boesch, C. (2012). Effect of two beta-alanine dosing protocols on muscle carnosine synthesis and washout. Amino Acids, 42(6), 2461-2472. doi:10.1007/s00726-011-1054-4
Stellingwerff, T., Bovim, I. M., & Whitfield, J. (2019). Contemporary Nutrition Interventions to Optimize Performance in Middle-Distance Runners. International journal of sport nutrition and exercise metabolism, 29(2), 106-116. doi:10.1123/ijsnem.2018-0241
Stellingwerff, T., Decombaz, J., Harris, R. C., & Boesch, C. (2012). Optimizing human in vivo dosing and delivery of beta-alanine supplements for muscle carnosine synthesis. Amino Acids, 43(1), 57-65. doi:10.1007/s00726-012-1245-7
Sweeney, K. M., Wright, G. A., Glenn Brice, A., & Doberstein, S. T. (2010). The effect of beta-alanine supplementation on power performance during repeated sprint activity. Journal of strength and conditioning research / National Strength & Conditioning Association, 24(1), 79-87. doi:10.1519/JSC.0b013e3181c63bd5
Van Thienen, R., Van Proeyen, K., Vanden Eynde, B., Puype, J., Lefere, T., & Hespel, P. (2009). Beta-alanine improves sprint performance in endurance cycling. Medicine and science in sports and exercise, 41(4), 898-903. doi:10.1249/MSS.0b013e31818db708
Varanoske, A. N., Hoffman, J. R., Church, D. D., Coker, N. A., Baker, K. M., Dodd, S. J., . . . Stout, J. R. (2019). Comparison of sustained-release and rapid-release beta-alanine formulations on changes in skeletal muscle carnosine and histidine content and isometric performance following a muscle-damaging protocol. Amino Acids, 51(1), 49-60. doi:10.1007/s00726-018-2609-4
Weyand, P. G., Sandell, R. F., Prime, D. N., & Bundle, M. W. (2010). The biological limits to running speed are imposed from the ground up. J Appl Physiol (1985), 108(4), 950-961. doi:10.1152/japplphysiol.00947.2009
Baixar

Distribuir



